Griefahn, B.
Zusammenfassung
Problemstellung: Da Morgentypen sich kaum an Nachtarbeit anpassen, war grundsätzlich zu klären, ob sich deren Phasenlage ebenso wie bei Neutral- und Abendtypen durch gezielte Lichtbehandlung verschieben lässt.
Methoden: 32 junge Männer (10 Morgen-, 11 Neutral-, 11 Abendtypen) absolvierten einen 40-Stunden-Versuch, der zwei aufeinander folgende Nächte umfasste. Abgesehen von zwei kurzen Unterbrechungen am zweiten Tag hielten sie bei einer Beleuchtung von 30 lux Bettruhe ein. Zur Bestimmung der Melatonin- und der Cortisolprofile wurden stündlich Speichelproben genommen. In der ersten Nacht erfolgte eine vierstündige Lichtbehandlung (1500 lux), die kurz nach dem erwarteten Beginn der individuellen Melatoninsynthese einsetzte.
Ergebnis: Die Lichtbehandlung verzögerte in der zweiten Nacht den Melatonin-Onset um 108, den Cortisol-Offset um 47 und den Cortisol-Onset um 110 Minuten und bewirkte so eine Verlängerung der Cortisolruhephase. Im Vergleich zu den Neutral- und Abendtypen war die Verschiebung des Melatonin-Onsets bei Morgentypen geringer, die Cortisolruhephase war verkürzt und die Cortisolproduktion tendenziell erhöht.
Schlussfolgerung: Morgentypen bleiben mutmaßlich selbst bei individuell terminierter Lichtbehandlung bezüglich der Anpassung an Nachtarbeit benachteiligt. Auf Grund erhöhter Cortisolspiegel ist nicht auszuschließen, dass sie bei wiederholter Nachtarbeit ein höheres Risiko zur Entwicklung kardiovaskulärer Erkrankungen haben.
Schlüsselwörter: Lichtexposition zirkadiane Phasenlage Chronotyp Cortisol Melatonin
Summary
Objective: As morning types scarcely adapt to night work this study tested the hypothesis that their phase position can be shifted by moderate bright light as well as in neither and in evening types.
Methods: 32 young men (10 morning-, 11 neutral, 11 evening types) took part in a 40-hours experiment over two consecutive nights. Apart from two short breaks on the second day they kept a strict bedrest at an illumination level of 30 lux. A 4-hours light pulse of 1500 lux was administered during the first night shortly after the expected onset of the individual melatonin synthesis. Saliva samples were taken hourly to determine melatonin and cortisol profiles.
Results: Light treatment caused in the second night a delay of the melatonin onset by 108, the cortisol offset by 47 and the cortisol onset by 110 minutes with a prolongation of the cortisol quiescent period. As compared to neither and to evening types the melatonin onset was less delayed in morning types, their cortisol quiescent period was shortened and their cortisol production elevated.
Conclusions: The results led to the hypothesis, that morning types remain disadvantaged even when treated with individually scheduled light where the elevated cortisol levels might, when these persons repeatedly work during the night, contribute to the genesis of cardiovascular diseases in this group.
Key words: light exposure circadian phase position diurnal type cortisol melatonin
Einleitung
Der Chronotyp (die individuelle Morgenorientierung) bezeichnet ein Persönlichkeitsmerkmal, das durch Abweichungen der individuellen zirkadianen Phasenlage vom 24-Stunden-Rhythmus des geophysikalischen Hell-Dunkelwechsels charakterisiert ist. Personen mit extremer Phasenlage, sog. Morgen- und Abendtypen, unterscheiden sich bezüglich der zeitlichen Variation ihres Verhaltens, ihrer Leistung, ihrer physiologischen Funktionen sowie in ihrer Fähigkeit, Schichtarbeit zu leisten. Während Abendtypen Probleme mit sehr früh beginnenden Schichten haben, können sich Morgentypen nicht an Nachtarbeit anpassen; ihre physiologischen Rhythmen bleiben dann dissoziiert und sie schlafen am Tage nach der Nachtschicht deutlich schlechter als Abendtypen10.
Die im zentralen Schrittmacher generierte zirkadiane Rhythmik wird insbesondere durch den zyklischen Hell-Dunkelwechsel auf den 24-Stunden-Rhythmus des geophysikalischen Tages synchronisiert, wobei das Hormon Melatonin mutmaßlich eine vermittelnde Rolle spielt. Licht verursacht akut eine Suppression der Melatoninsynthese, der eine Verschiebung des zirkadianen Systems folgt. Ausmaß und Richtung der Phasenverschiebung sind durch den Zeitpunkt der Lichteinwirkung bestimmt. Abends appliziertes Licht bewirkt eine Verzögerung, morgens einwirkendes Licht eine Voreilung. Dieser systemische, zahlreiche physiologische Funktionen umfassende chronobiologische Effekt wird auch durch artifizielles Licht erzielt. Deshalb ist die Annahme plausibel, dass zu definierten Abschnitten des individuellen zirkadianen Rhythmus appliziertes Licht (zum Beispiel kurz nach Einsetzen der Melatoninsynthese) unabhängig von der jeweiligen Phasenlage eine jeweils gleiche Verschiebung verursacht, dass Morgentypen sich also bei entsprechender Behandlung ebenso an Nacht-arbeit anpassen wie Abendtypen.
Dies war in der hier beschriebenen Pilotstudie grundsätzlich zu prüfen. Dazu wurden 32 junge Männer unterschiedlicher Phasenlagen in einem 40-Stunden-Versuch über zwei aufeinander folgende Nächte beobachtet. In der ersten Nacht wurde kurz nach dem erwarteten individuellen Melatonin-Onset 4 Stunden lang helles Licht appliziert, um eine zirkadiane Verzögerung in der zweiten Nacht zu erzielen.
Probanden
Die Probanden gaben nach entsprechender Aufklärung ihr schriftliches Einverständnis zu der von der Ethikkommission genehmigten Untersuchung. 47 junge Männer (1834 Jahre), bei denen die subjektive Phasenlage mit dem Morgen-Abend-Fragebogen [D-MEQ6] ermittelt worden war, absolvierten eine modifizierte Constant Routine CR0 zur Bestimmung des individuellen Melatoninprofils und damit des Zeitraums für die individuelle Lichtbehandlung, die kurz nach Beginn der Melatoninsynthese einsetzen sollte.
Constant Routine (CR0)
Die CR0 wurde ebenso wie der 40-Stunden-Versuch in separaten schallisolierten Räumen durchgeführt. Während der CR0 hielten die Probanden von 189 Uhr Bettruhe ein. Da sich die Melatoninkonzentration mit der Körperhaltung ändert und erst nach etwa 20 Minuten ein steady state erreicht, durfte das Bett für gelegentliche Toilettengänge nur innerhalb der auf die stündliche Speichelprobennahme folgenden 20 Minuten verlassen werden. In allen Räumen, zu denen die Probanden Zugang hatten (Flure, Badezimmer, im 40-Stunden-Versuch auch der Speiseraum) wurde die Beleuchtung auf 30 lux eingestellt. Die Probanden wurden kontinuierlich über ein Videosystem beobachtet; sie durften lesen, schreiben und zwischen den Probennahmen schlafen.
Experimente zur Licht induzierten Phasenverschiebung
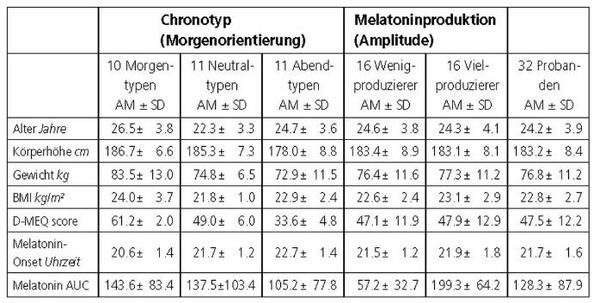
Basierend auf den Ergebnissen der CR0 und der D-MEQ-Scores wurden 10 Morgen-, 11 Neutral- und 11 Abendtypen, deren Daten in Tabelle 1 aufgelistet sind, für den 40-Stunden-Versuch ausgewählt. Deren zirkadiane Amplitude wurde, indiziert durch die Melatoninmenge, bei jeweils 16 Personen als hoch bzw. niedrig eingestuft (niedrig: kein Melatoninwert >15 pg/ml). Die experimentellen Untersuchungen liefen nach einem von van Cauter et al.13 entwickelten Design ab. Der Versuch begann um 17 Uhr und war um 9 Uhr des übernächsten Tages beendet. Abgesehen von zwei kurzen Unterbrechungen am zweiten Tag (1012 Uhr, 1617 Uhr, Duschen, Mahlzeit) hielten die Probanden Bettruhe ein. Die Beleuchtung war auf 30 lux eingestellt; die jeweils 4-stündige Lichtbehandlung (1500 lux) setzte entsprechend des in der CR0 bestimmten Melatoninprofils in der ersten Nacht (N1) zu der auf den erwarteten Melatonin-Onset folgenden vollen Stunde ein. Ein Kontrollversuch war nicht vorgesehen, da van Cauter et al.13 in ihrem Kontrollexperiment keine Veränderung der zirkadianen Phasenlage beobachteten und da die Reproduzierbarkeit des Melatoninprofils intraindividuell sehr hoch ist1.
Lichtexposition
Die Lichtquelle bestand aus mehreren, die Betten umgebenden Leuchtstoffröhren (OSRAM DeLuxe 1158W/12, Lumilux de Luxe, Tageslicht 12950), deren weißes Licht gegen die weiß gestrichenen Decken, Wände und Fußböden gerichtet war, was eine gleichmäßige Beleuchtung (von 1500 lux) in allen Blickrichtungen gewährleistete.
Speichelprobennahmen zur Bestimmung der Hormonprofile
erfolgten stündlich mit einem Watteröllchen (Salivette®), das nach dem Einspeicheln zentrifugiert wurde. Der Speichel wurde bei 20°C bis zur Analyse eingefroren. Die Melatoninkonzentration wurde mittels eines kommerziellen, kompetitiven Radioimmunoassays (RIA) mit einer Detektionsgrenze bei ca. 0.8 pg/ml Speichel, die Cortisolkonzentration mit einem Lumineszenz-Immumoassay (LIA, IBL) mit einer Nachweisgrenze von ca. 0.16 ng/ml Speichel bestimmt.
Auswertung, Statistik
Als Melatonin-Onset wurde jeder visuell erkennbare Anstieg vor der Lichtapplikation eingestuft. Bei 7 Probanden trat dieser wegen unerwartet starker zeitlicher Variabilität der Melatoninproduktion jedoch nicht vor der Lichtapplikation ein, weshalb das Cortisol-Profil als weiterer Indikator der zirkadianen Rhythmik bestimmt wurde. Offset und Onset der Cortisolproduktion (Beginn und Ende der Cortisolruhephase) waren definiert als die Zeitpunkte, zu denen 60% des über die Versuchsdauer errechneten arithmetischen Mittels unter- bzw. überschritten wurden (was dem von mehreren Autoren verwendeten 50%-Wert des über 24 Stunden errechneten arithmetischen Mittels entspricht). Damit ließen sich Beginn und Ende der Cortisolruhephase für 22 bzw. 29 Probanden ermitteln. Alle Berechnungen wurden mit der größtmöglichen Anzahl von Daten vorgenommen, Korrelationen aber nur für die 17 Probanden mit vollständigem Datensatz (Melatonin-Onset, Cortisol-Offset und Cortisol-Onset) errechnet.
Der Chronotyp war durch den D-MEQ-Score indiziert, die als Confounder berücksichtigte zirkadiane Amplitude durch die mit dem Maximum hoch korrelierende Fläche unter der Melatoninkurve (area under curve, AUC). Die Verschiebung der Hormonprofile wurde durch die Differenz der in beiden Nächten erfassten zeitlichen Parameter bestimmt (N2 N1, Melatonin-Onset, Cortisol-Offset, Cortisol-Onset). Statistische Berechnungen wurden mit dem Generalisierten Linearen Modell (GLM), dem Wilcoxon Test und Pearson Korrelationen vorgenommen.
Ergebnisse
Constant Routine (CR0)
Abgesehen von den Einteilungskriterien (Phasenlage, Amplitude) bestanden keine signifikanten Unterschiede zwischen den einzelnen Untergruppen (Tab. 1).
Die subjektive Phasenlage (D-MEQ-Score) korrelierte signifikant mit dem Melatonin-Onset, der zwischen 18:48 und 00:48 Uhr variierte (r = 0.47, p0.01), nicht jedoch mit der zwischen 20314 pg/ml*h variierenden zirkadianen Amplitude (AUC).
Lichtinduzierte Phasenverschiebung
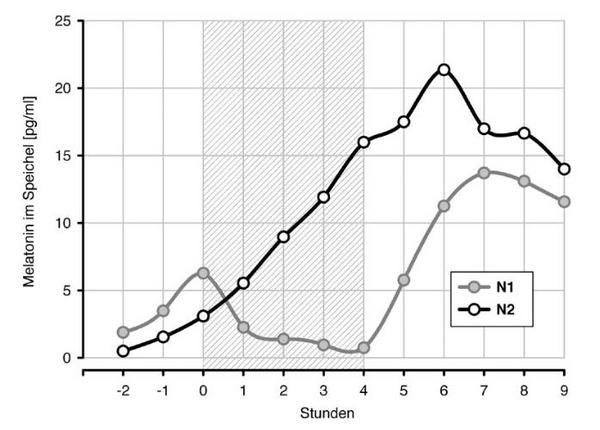
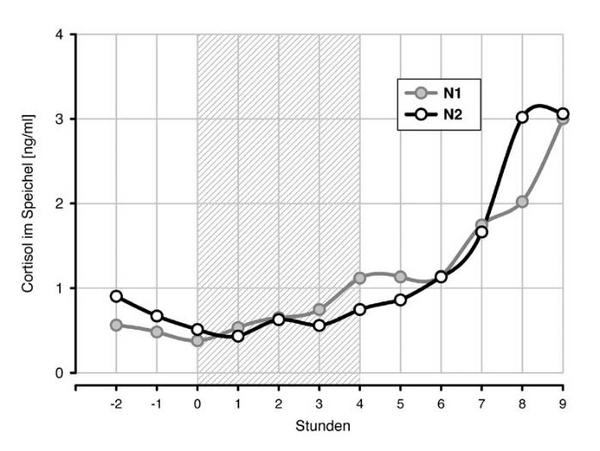
Abb. 1a und 1b zeigen die Verläufe der Melatonin- und der Cortisolkonzentrationen, synchronisiert über den individuellen Beginn der Lichtapplikation. Trotz der eher moderaten Beleuchtung von 1500 lux, bewirkte diese bei 31 von 32 Probanden eine vollständige Unterdrückung der Melatoninsynthese (Abb. 1a) und eine deutliche Verzögerung des Melatonin-Onsets in der zweiten Nacht. Die Cortisolkonzentration (Abb. 1b) war durch die Lichteinwirkung nicht beeinflusst, zeigte in der zweiten Nacht aber ebenfalls eine Verzögerung (Rechtsverschiebung) gegenüber der ersten Nacht.
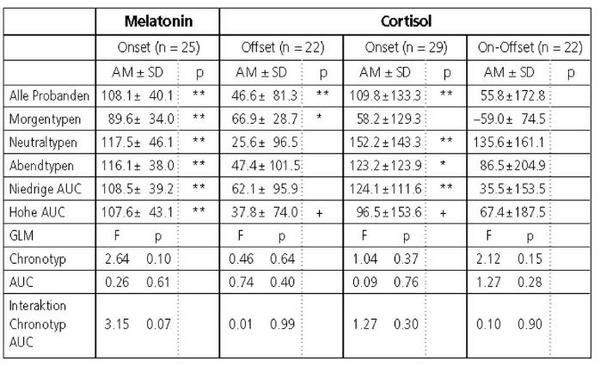
Die Verzögerung der zeitlichen Parameter der Hormonprofile ist in Tabelle 2 quantifiziert. Sie betrug für den Melatonin-Onset 108, für den Offset und den Onset des Cortisols 47 bzw. 110 Minuten (p 0.01). Die um eine Stunde stärkere Verschiebung der Onsets von Melatonin und Cortisol gegenüber dem Cortisol-Offset indiziert sowohl eine partielle Dissoziation zwischen beiden Hormonprofilen als auch eine Distorsion des Cortisolprofils mit einer Verlängerung der Ruhephase.
Chronotyp
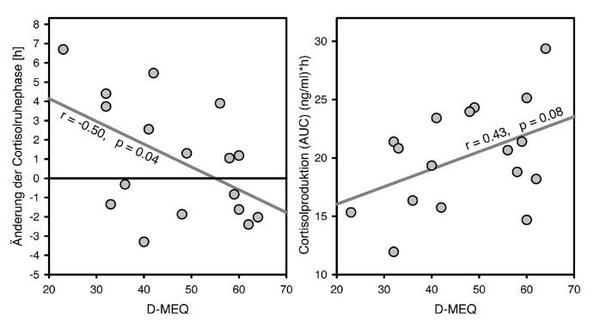
Die Verzögerung des Melatonin-Onsets in der zweiten Nacht war bei allen Chronotypen zwar signifikant (p 0.01), bei den Morgentypen jedoch um 28 Minuten geringer als bei den Neutral- und den Abendtypen. Im Vergleich zu letzteren war der Cortisol-Offset nur bei den Morgentypen signifikant verzögert, der Cortisol-Onset hingegen nicht. Daraus resultierte nur in dieser Untergruppe eine Verkürzung der Cortisolruhephase, während sie in den anderen Gruppen verlängert war (Abb. 2).
Das GLM ergab, abgesehen von einem möglichen Einfluss des Chronotyps auf die Verzögerung des Melatoninprofils (Tab. 2), keinen signifikanten Einfluss des Chronotyps oder der zirkadianen Amplitude auf die Verzögerung der Phasenlage.
Korrelationen
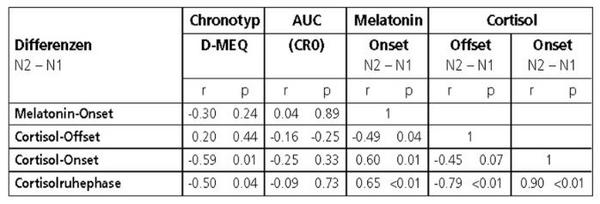
Nach Tabelle 3 war die Verzögerung des Melatonin-Onsets invers mit der des Cortisol-Offsets (r = 0.49, p = 0.04), aber direkt mit den Cortisol-Onsets (r = 0.60, p 0.01) und so mit einer zunehmenden Verlängerung der Cortisolruhephase verknüpft (r = 0.65, p 0.01). Desgleichen war, wie Abbildung 2 zeigt, die subjektive Morgenorientierung signifikant aber invers mit der Verlängerung der Cortisolruhephase (r = 0.50, p = 0.04) korreliert, während die Cortisolproduktion in der zweiten Nacht tendenziell mit der Morgenorientierung zunahm (r = 0.43, p= 0.08).
Verzögerung der Hormonprofile
Anders als in Untersuchungen anderer Autoren führte die hier eingestellte eher moderate Beleuchtung von 1500 lux bei 31 von insgesamt 32 Probanden zu einer vollständigen Suppression der Melatoninsynthese9, 14. Dies wurde durch die Anordnung der Lichtquelle erzielt, bei der in jeder Blickrichtung die gleiche Beleuchtung bestand, wozu die weißen Decken, Wände, Fußböden und die weiße Bettwäsche beitrugen.
Ausmaß und Richtung lichtinduzierter Verschiebungen der Phasenlage hängen im Wesentlichen von der zeitlichen Distanz der Lichteinwirkung zum Minimum der Körperkerntemperatur (12h vor dem Melatonin-Maximum) und von der Beleuchtungsstärke ab2. Zeitzer et al.14 stellten die Beziehung der Melatoninsuppression und der Verschiebung des Melatoninprofils in Abhängigkeit zur Beleuchtung mittels logistischer Regressionen dar. Da die vollständige Melatoninsuppression schneller erreicht ist als die maximale Verschiebung, lässt sich letztere nicht aus der Melatoninsuppression vorhersagen. Die Komplexität des Zusammenhangs kommt unter anderem dadurch zum Ausdruck, dass die hier beobachtete Verzögerung trotz vollständiger Melatoninsuppression um 0.6 Stunden geringer war als sie von Dawson et al.5 mit einer einzigen 4-stündigen Lichtapplikation von 10 000 12 000 lux (also vollständiger Melatoninsuppression) erzielt worden war. Sie war andererseits um 0.6 Stunden größer als inden Untersuchungen von van Cauter et al.13, die eine Beleuchtung von 5000 lux 3 Stunden lang einwirken ließen.
Zeitliche Beziehung zwischen Melatonin- und Cortisolprofilen
Unter normalen Bedingungen besteht zwischen beiden Hormonprofilen eine enge zeitliche Beziehung, die sogar nach lichtinduzierter Verschiebung des zirkadianen Systems und selbst nach mehrwöchiger Entkopplung der zirkadianen Rhythmen und des Schlaf-Wachwechsels stabil bleibt4, 12. Die Stabilität dieser zeitlichen Beziehungen zeigt sich in den hier vorgenommenen Untersuchungen in der nahezu gleichen und signifikant miteinander korrelierenden Verzögerung des Onsets von Melatonin und Cortisol, wird andererseits aber durch die um eine Stunde geringere Verschiebung des Cortisol-Offsets in Frage gestellt. Die unterschiedliche zeitliche Verzögerung von Cortisol-Onset und Cortisol-Offset bewirkte eine Verlängerung der Cortisolruhephase.
Caufriez et al.3, die den Ruhe-Aktivitäts-Zyklus um 8 Stunden vorverlagert hatten, fanden hingegen durch selektives Voreilen des Cortisol-Onsets eine Verkürzung der Cortisolruhephase.
Individuelle zirkadiane Phasenlage (Chronotyp, Morgenorientierung)
Die Annahme, dass die auf das individuelle Melatoninprofil terminierte Lichtapplikation zur jeweils gleichen Verschiebung der zirkadianen Phasenlage führt, wurde hier nicht bestätigt. Statt dessen war die Phasenlage, gemessen am Melatonin-Onset, bei den Morgentypen um 28 Minuten weniger verschoben als bei Neutral- und Abendtypen; die Cortisolruhephase war nur in dieser Untergruppe verkürzt und deren Cortisolproduktion in der zweiten Nacht tendenziell erhöht.
Da eine erhöhte Cortisolproduktion in der Cortisolruhephase im Alter typischerweise erhöht ist, vermuten Caufriez et al.3, dass wiederholte Änderungen des Zeitregimes, wie etwa bei Schichtarbeit zur Entwicklung alterstypischer metabolischer und kognitiver Defizite beitragen können. Darüber hinaus ist hinreichend gesichert, dass erhöhte Cortisolspiegel zur Genese kardiovaskulärer Erkrankungen beitragen und dass diese Erkrankungen bei Schichtarbeitern gehäuft auftreten7, 8. Aus den Ergebnissen der vorliegenden Arbeit lässt sich die Hypothese ableiten, dass die Probleme von Morgentypen, sich an Nachtarbeit anzupassen, endogener Natur sind und Morgentypen, sofern die beobachteten Veränderungen bei langfristiger Nachtarbeit wiederholt auftreten, ein höheres Risiko haben, kardiovaskuläre Erkrankungen zu entwickeln.
Amplitude der zirkadianen Rhythmik (AUC)
Die zirkadiane Amplitude wurde hier als möglicher Confounder berücksichtigt, da Reinberg et al.11 bei Personen mit geringer zirkadianer Amplitude eine schnellere Anpassung an Nachtarbeit fanden. In den hier dargestellten Untersuchungen hatte die Amplitude, indiziert durch die gesamte Melatoninproduktion (AUC) jedoch keine Bedeutung für das Ausmaß der lichtinduzierten Verschiebung der Phasenlage.
Schlussfolgerungen
Die vorliegende Untersuchung prüfte die Hypothese, dass eine individuell auf den Beginn der Melatoninsynthese terminierte Lichtbehandlung das zirkadiane System von Personen unterschiedlicher zirkadianer Phasenlage gleichermaßen verzögert. Dies wurde nicht voll bestätigt. Im Vergleich zu Neutral- und Abendtypen war die lichtinduzierte Verzögerung der Hormonprofile bei Morgentypen geringer und deren Cortisolruhephase verkürzt.
Da diese Untersuchung nicht auf die Bestimmung des Cortisols ausgerichtet war, sind die Ergebnisse zurückhaltend zu interpretieren. Sie lassen aber vermuten, dass Morgentypen mutmaßlich selbst bei individueller Lichtbehandlung benachteiligt bleiben und dass diese, sofern die Verkürzung der Cortisolruhephase bei häufiger Nachtarbeit wiederholt auftritt, ein erhöhtes Risiko zur Entwicklung kardiovaskulärer Erkrankungen haben.
Die Untersuchung wurde durch die Deutsche Forschungsgemeinschaft finanziert.
Literatur
1 Arendt J. Melatonin and the pineal gland: influence on mammalian seasonal and circadian physiology. Rev Rep 1998; 3: 1322
2 Boivin DB, Czeisler CA Resetting of circadian melatonin and cortisol rhythms by ordinary room light. Neuroendocrinol 1998; 9: 779782
3 Caufriez A, Moreno-Reyes R, Leproult R, Vertongen F, van Cauter E, Copinschi G. Immediate effects of an 8-h advance shift of the rest-activity cycle on 24-h profiles of cortisol. Am J Physiol Endocrinol Metab 2002; 282: E1147E1153
4 Czeisler CA, Duffy JF, Shanahan TL, Brown EN, Mitchell JF, Rimmer DW, Ronda JM, Silva EJ, Allan JS, Emens JS, Dijk DJ, Kronauer RE Stability, precision, and near-24-hour period of the human circadian pacemaker. Science 1999; 284: 21772181
5 Dawson D, Lack L, Morris M. Phase resetting of the human circadian pacemaker with use of a single pulse of bright light. Chronobiol Int 1993; 10: 94102
6 Griefahn B, Künemund C, Bröde P, Mehnert P. Zur Validität der deutschen Übersetzung des Morningness-Eveningness-Questionnaires von Horne und Östberg. Somnologie 2001; 5: 7180
7 Knutsson A. Methodological aspects of shift-work research. Chronobiol Int 2004; 21: 10371047
8 Knutsson A, Bøggild H. Shiftwork and cardiovascular disease: Review of disease mechanisms. Rev Environ Health 2000; 15: 359372
9 Lewy AJ, Wehr TA, Goodwin FK. Light suppresses melatonin secretion in humans. Science 1980; 210: 12671269
10 Moog R. Individuelle circadiane Phasenlage ein Prädiktor der Nacht- und Schichtarbeitstoleranz. Inaugural-Diss. Univ. Marburg, 1988
11 Reinberg A, Motohashi Y, Bourdeleau P, Andlauer P, Lévi F, Bicakova-Rocher A. Alteration of period and amplitude of circadian rhythms in shift workers. Eur J Appl Physiol 1988; 57: 1525
12 Selmaoui B, Touitou Y. Reproducibility of the circadian rhythms of serum cortisol and melatonin in healthy subjects: a study of three different 24-h cycles over six weeks. Life Sci 2003; 73: 33393349
13 van Cauter E, Sturis J, Byrne MM, Blackman JD, Leproult R, Ofek G, L”Hermite-Balériaux M, Refetoff S, Turek FW, van Reeth O. Demonstration of rapid light-induced advances and delays of the human circadian clock using hormonal phase markers. Am J Physiol 1994; E953E963
14 Zeitzer JM, Dijk DJ, Kronauer RE, Brown EN, Czeisler CA. Sensitivity of the human circadian pacemaker to nocturnal light: melatonin phase resetting and suppression J Physiol (Lond) 2000; 526: 695702
Anschrift der Verfasserin:
Univ.-Prof. Dr. Barbara Griefahn
Institut für Arbeitsphysiologie
an der Universität Dortmund
Ardeystr. 67, D-44139 Dortmund
Tel. +49 231 1084 221, Fax +49 231 1084 400
email: griefahn@ifado.de


